Какие факторы стимулируют эритропоэз
Стадии дифференцировки красных клеток крови. Первой клеткой, которую можно идентифицировать как принадлежащую к группе красных клеток крови, является проэритробласт, показанный как отправная точка на рисунке ниже.
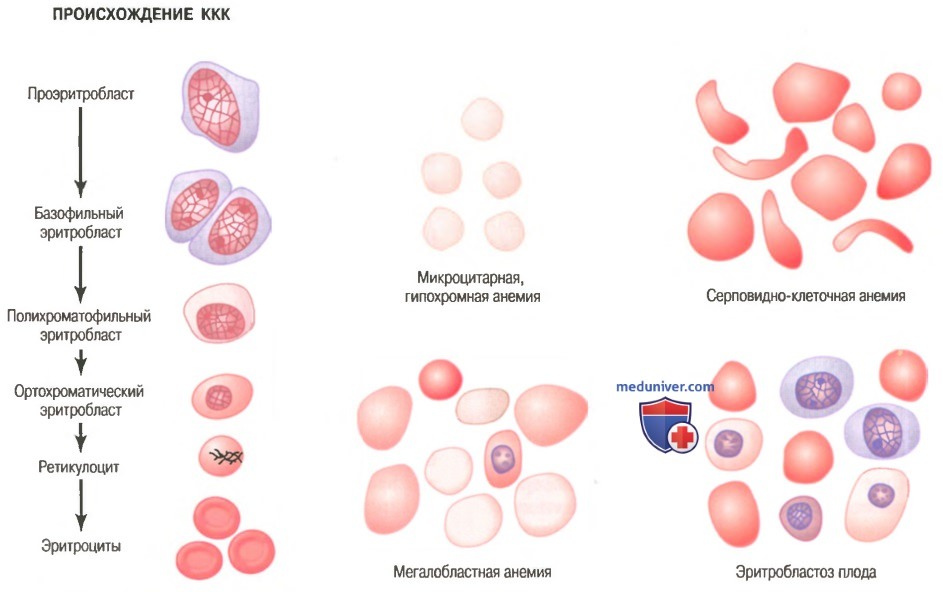 Происхождение нормальных красных клеток крови (ККК) и их особенности при разных типах анемий
Происхождение нормальных красных клеток крови (ККК) и их особенности при разных типах анемий
При соответствующей стимуляции большие количества этих клеток формируются из стволовых клеток КОЕ-Э.
Сразу после образования проэритробласт делится много раз, в результате формируются много зрелых красных клеток крови. Клетки первой генерации называют базофилъными эритробластами, поскольку они окрашиваются основными красителями; к этому времени клетка накапливает очень мало гемоглобина.
В последующих генерациях, как показано на рисунке, клетки наполняются гемоглобином до концентрации, равной примерно 34%, ядро сжимается до небольшого размера, и его конечный остаток абсорбируется или выдавливается из клетки. В то же время абсорбируется эндоплазматический ретикулум. На этом этапе клетку называют ретикулоцитому поскольку она все еще содержит небольшое количество базофильного материала, состоящего из остатков аппарата Гольджи, митохондрий и некоторых других цитоплазматических органелл.
Во время этой ретикулоцитарной стадии клетки выходят из костного мозга в кровеносные капилляры путем диапедеза (просачиваясь сквозь поры капиллярной мембраны).
Остаточный базофильный материал ретикулоцита в норме исчезает в течение 1-2 сут, и после этого клетка становится зрелым эритроцитом. В связи с короткой жизнью ретикулоцитов их концентрация среди всех клеток крови обычно немного ниже 1%.
Регуляция продукции красных клеток крови. Роль эритропоэтина
Общая масса эритроцитов в системе кровообращения регулируется в узких пределах, поэтому, с одной стороны, (1) всегда имеется адекватное количество красных клеток крови для обеспечения достаточного транспорта кислорода из легких к тканям, с другой стороны, (2) клеток не становится так много, чтобы они мешали кровотоку. Известные в настоящее время данные об этом контроле представлены в виде схемы на рисунке ниже и заключаются в следующем.
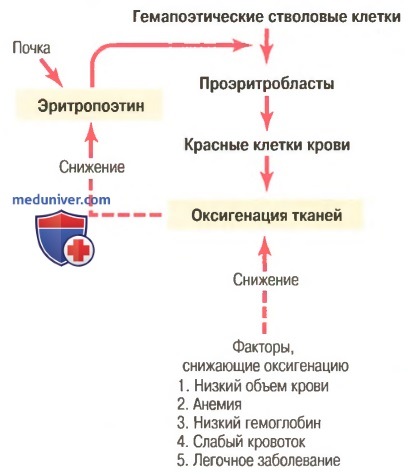 Увеличение продукции красных клеток крови, связанное с функцией механизма эритропоэтина при снижении оксигенации тканей
Увеличение продукции красных клеток крови, связанное с функцией механизма эритропоэтина при снижении оксигенации тканей
а) Оксигенация тканей является наиболее важным регулятором продукции красных клеток крови. Любое состояние, ведущее к уменьшению доставки кислорода тканям, обычно увеличивает скорость образования эритроцитов. Следовательно, если в результате кровотечения или любого другого состояния у человека развивается анемия, костный мозг немедленно усиливает продукцию красных клеток крови. Кроме того, разрушение значительной части костного мозга по любой причине, особенно при рентгенотерапии, ведет к гиперплазии оставшегося костного мозга, что отражает попытку удовлетворить потребности организма в эритроцитах.
На очень больших высотах, где количество кислорода в воздухе сильно снижено, снабжение тканей кислородом становится недостаточным, и продукция красных клеток крови резко возрастает. В этом случае образование эритроцитов регулирует не их концентрация в крови, а несоответствие между доставкой кислорода тканям и потребностями тканей в кислороде.
Скорость продукции красных клеток крови возрастает также при различных болезнях системы кровообращения, сопровождаемых снижением кровотока через периферические сосуды, особенно на фоне недостаточности поглощения кислорода при прохождении крови через легкие. Так, тканевая гипоксия, характерная для хронической сердечной недостаточности и многих легочных заболеваний, стимулирует продукцию красных клеток крови, что ведет к увеличению гематокрита и, как правило, общего объема крови.
б) Эритропоэтин стимулирует продукцию красных клеток крови, а его образование возрастает в ответ на гипоксию. Главным стимулом для продукции эритроцитов при состояниях с низким содержанием кислорода является циркулирующий в крови гормон эритропоэтин — гликопротеин с молекулярной массой около 34000. При отсутствии эритропоэтина гипоксия практически не стимулирует продукцию эритроцитов. Но когда система эритропоэтина функционирует, гипоксия вызывает заметное увеличение секреции эритропоэтина, а он, в свою очередь, усиливает образование красных клеток крови до тех пор, пока гипоксия не исчезнет.
в) Роль почек в формировании эритропоэтина. У здорового человека примерно 90% всего эритропоэтина синтезируется в почках; остальная часть образуется в основном в печени. Пока точно не известно, где именно в почках синтезируется эритропоэтин. Одним из возможных мест секреции эритропоэтина являются эпителиальные клетки почечных канальцев, поскольку анемичная кровь околоканальцевых капилляров не способна удовлетворить их высокую потребность в кислороде, что стимулирует продукцию эритропоэтина.
Иногда секрецию эритропоэтина почками стимулирует гипоксия не в почках, а в других частях тела, что позволяет предположить возможное существование некоего внепочечного сенсора, который посылает к почкам дополнительный сигнал, стимулирующий продукцию этого гормона. В частности, норадреналин, адреналин и некоторые из простагландинов стимулируют продукцию эритропоэтина.
Когда у человека удаляют обе почки или если они разрушены болезнью, у человека неминуемо развивается тяжелая анемия, поскольку эритропоэтин, синтезируемый другими тканями (в основном печенью), составляет лишь 10% нормы и может обеспечить формирование не более половины красных клеток крови, необходимых организму.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
— Вернуться в оглавление раздела «Физиология человека.»
Стимуляторы эритропоэза ( по материалам статьи David Goldsmith )

Первоначальный энтузиазм в отношении этого нового подхода почти полностью основывался на демонстрации предотвращения переливания крови и на ограниченных наборах данных, показывающих улучшение толерантности к физической нагрузке, пройденного расстояния, качества жизни и улучшение размеров и производительности левого желудочка. Профиль побочных эффектов искусственной инженерии более высокого гематокрита с использованием эпоэтина был очевиден с самого начала.
Вероятно, лечение пациентов с хроническим заболеванием почек эпоэтинами только для «коррекции» уровня гемоглобина или для соблюдения рекомендаций не является хорошим лекарством, так же как и решение о необходимости диализа для пациента на основе исключительно измерений СКФ. Некоторые исследователи утверждает, что нижний предел для гемоглобина потенциально может быть установлен на таком уровне, который не приводит к заметному увеличению связанной с переливанием аллосенсибилизации. Конечно, эпоэтины не отменяют, а значительно снижают показатели переливания крови. Тем не менее, некоторые оговорки должны быть введены. Во-первых, влияние лейкодеплированной крови на иммунологическую систему является комплексным: донор-специфическое переливание или одно несоответствующее переливание HLA-DR может быть безвредным или полезным, и этот эффект может варьироваться между мужчинами и женщинами. Беременность / аборт остаются самым большим фактором риска аллосенсибилизации. Во-вторых, в Великобритании только
50% пациентов, находящихся на диализе, попадают в лист ожидания по пересадке почки, и из них только 25% или около того перенесли пересадку через 2 года после включения в список ожидания.
Регуляция кроветворения: механизмы и важные факторы
» data-image-caption=»» data-medium-file=»https://unclinic.ru/wp-content/uploads/2019/05/funkcii-krovetvorenija.jpg» data-large-file=»https://unclinic.ru/wp-content/uploads/2019/05/funkcii-krovetvorenija.jpg» title=»Регуляция кроветворения: механизмы и важные факторы»>
Алена Герасимова (Dalles) Разработчик сайта, редактор
Постоянство клеточного состава крови, его обновление осуществляются благодаря взаимосвязи крови и органов, образующих ее элементы (кроветворных).
В костном мозге созревают красные кровяные тельца, зернистые лейкоциты и тромбоциты. Общий вес его у взрослого человека приблизительно составляет 1500 г. Лимфатические узлы, селезенка образуют лимфоциты и моноциты.
Особенности процесса образования клеток крови: теории и факты
Процесс образования клеток крови идет непрерывно в течение всей человеческой жизни, интенсивность его строго соответствует потребностям организма.
По одной из современных теорий следует, что клетки крови человека — эритроциты, лейкоциты и тромбоциты происходят из единой родоначальной материнской клетки, так называемой «стволовой». Путем ее деления и развития появляются клепки, предопределяющие различные ветви кроветворения: образование эритроцитов, зернистых лейкоцитов (гранулоцитов), незернистых лейкоцитов (агранулоцитов), тромбоцитов.
Порожденная общей «стволовой» клеткой, каждая из этих ветвей имеет и свою собственную родоначальную клетку. В процессе деления и постепенного созревания и преобразования этих костномозговых элементов появляются зрелые клетки, поступающие в кровь.
К чести русской науки следует оказать, что мысль о происхождении всех клеток крови из единого источника принадлежит знаменитому русскому ученому-гистологу А. А. Максимову, создавшему еще в 1900—1914 гг. свою теорию кроветворения. Эти исследования нашли подтверждение и дальнейшее развитие в трудах советских исследователей.
Вместе с тем в некоторыми учеными высказывалась мысль о том, что еще до рождения человека в кроветворных органах предопределен вид каждой кроветворной линии — гранулоцитарной, эритроцитарной, лимфоцитарной. В пользу такой точки зрения приводятся данные биохимических исследований клеток крови и костного мозга.
Так, советские биохимики П. Ф. Сейц и П. С. Луганова обнаружили, что для определенных линий кроветворных элементов характерен определенный вид энергетического обмена. На основании этих данных они полагали, что и происхождение клеточных форм на каком-то этапе должно быть различным, поскольку характерный тип обмена (как группа крови, резус-фактор), возникший в клетке в начальном периоде ее развития, сохраняется во всех клеточных популяциях (производных данной линии).
Из всего сказанного можно сделать заключение о том, что кровь обладает многообразными функциями, имеющими первостепенное значение для существования организма. Всякое нарушение постоянства состава этой внутренней среды организма чревато далеко идущими последствиями, приводящими к нарушению здоровья человека.
Как осуществляется кроветворение: механизмы
Процессы разрушения красных кровяных шариков и их образования строго сбалансированы. Если организм теряет какое-то количество крови, то не проходит 2—3 недель, как снова восстанавливается исходный уровень числа эритроцитов и концентрации гемоглобина. При этом всегда наблюдается значительное убыстрение образования красных кровяных телец (эритропоэза) в костном мозге.
Не вызывает сомнений факт существования в организме особых механизмов регуляции эритропоэза, хорошо выявляемых тогда, когда под влиянием каких-либо причин резко уменьшается количество эритроцитов и в связи с этим развивается кислородное голодание — гипоксия.
Законно предположить, что уменьшение снабжения организма кислородом автоматически приводит к увеличению продукции красных кровяных телец.
Возникает вопрос о механизме «эритроцитостимулирующего» действия кислородного голодания. Большим количеством исследований установлено, что этот фактор убыстряет кроветворение через посредство особого вещества, стимулирующего эрицропоэз и получившего название «эритропоэтин».
В 1906 г. два французских исследователя — Карно и Дефляндер — обнаружили, что сыворотка крови, взятая у кроликов через 20 часов после массивной кровопотери и введенная другому здоровому кролику, способствовала у последнего приросту эритроцитов на 2—3 млн. в 1 мм3 крови, а также увеличению количества гемоглобина.
Последующие эксперименты показали, что кислородная недостаточность любого происхождения способна повышать эритростимулирующие свойства кровяной сыворотки.
Наиболее убедительные доказательства существования в организме стимулятора красного кроветворения были представлены в опытах на искусственно сращенных между собой (наподобие сиамских близнецов) крысах.
Этот интересный опыт выглядел так: одна из крыс дышала газовой смесью, содержащей пониженное количество кислорода, а ее партнер — воздухом с нормальным содержанием кислорода. И оказалось, что у обоих животных в костном мозге происходило одинаковое разрастание клеток «красного ряда», а в периферической крови — значительное увеличение эритроцитов.
Объяснить это можно следующим образом: у крысы под влиянием кислородного голодания образуется вещество эритростимулирующего действия, т. е. эритропоэтин, который переходит с кровью через сращенные кровеносные сосуды в организм партнера и вызывает у него активизацию кроветворения.
В каком месте организма образуется эритропоэтин?
Многочисленные клинические наблюдения и особенно опыты на животных представили убедительные аргументы в пользу почечного происхождения эритропоэтина.
Было показано, что двустороннее удаление почек ликвидирует способность организма образовывать эритропоэтин в ответ на кровопотерю или на недостачу кислорода по другой причине. Последующая же подсадка почки, взятой от другого животного, вызывала очень быстрое восстановление эритропоэза в костном мозге.
Роль витамина В12 в кроветворении
В кроветворении принимают участие различные витамины, среди которых особая роль принадлежит витамину В12, содержащему кобальт.
Источником витамина В12 служат продукты животного происхождения; в растительных продуктах он отсутствует. Благодаря этому витамину поддерживается нормальный процесс созревания эритроцитов у здорового человека.
В сутки взрослому человеку необходимо 3—5 мг витамина В12. Как показали современные исследования, витамин В12, попавший в организм с пищей, всасывается в кишечнике лишь при соединении его с особым белком — гастромукопротеином (который иначе называется «внутренний фактор»).
Гастромукопротеин вырабатывается у человека железами желудка и обладает способностью образовывать с витамином В12 комплексное соединение. Оказалось, что этот белок предохраняет витамин от пожирания микроорганизмами, заселяющими кишечник. Таким образом, он выступает в роли «проводника» витамина В12 и спасает его от разрушающего действия микробов.
Всосавшийся витамин накапливается в печени и затем используется для целей кроветворения по мере необходимости.
Установлено, что витамин B12 принимает активное участие в образовании соединений, являющихся составными частями нуклеиновых кислот, — тех самых кислот, коими так богаты ядра клеток и которые определяют основные наследственные признаки организма.
В случае нехватки витамина B12 задерживается синтез нуклеиновых кислот, в результате чего неизбежно нарушается деление постоянно размножающихся кроветворных клеток. Тогда в костном мозге вместо нормальных эритробластов появляются огромные, медленно созревающие клетки, получившие название мегалобластов (от греческого слова «мегалос» — огромный).
На этой почве происходит развитие тяжелого малокровия — злокачественная анемия.
Роль гормонов и нервной системы в кроветворении
Вся сложная, необыкновенно подвижная система крови находится под постоянным влиянием эндокринной и нервной систем. Гормоны (от греческого слова «гормао» — возбуждаю), выделяемые эндокринными органами (железами внутренней секреции), попадают непосредственно в кровь.
Через нее гормоны осуществляют связь одних органов и систем с другими. Они оказывают регулирующее влияние на различные функции организма, в том числе и на кроветворение. Так воздействуют гормоны, вырабатываемые передней долей гипофиза, щитовидной железой, корой надпочечников, половыми железами.
Значительное влияние на процессы кроветворения и распределения элементов крови в сосудах и депо оказывает и, нервная система.
Какие факторы стимулируют эритропоэз
Эритропоэз и механизмы его регуляции
В эмбриональном периоде кроветворение осуществляется вначале в кровяных островках желточного мешка, затем примерно спустя 5 недель эмбрионального развития – в печени. Селезенка включается в процесс кроветворения с 16 недели внутриутробного развития. Первые гемопоэтические элементы появляются в костном мозге на 2-ом месяце эмбрионального развития, однако миелоидный период кроветворения начинается на 4-5-м месяцах эмбрионального развития, вытесняя постепенно кроветворение в печени и селезенке. Костномозговой эритропоэз осуществляется вне синусов, в строме костного мозга, то есть эктраваскуляторно. К моменту рождения ребенка костный мозг развивается полностью, а экстрамедуллярное кроветворение практически завершается. Постэмбриональный период кроветворения начинается после рождения ребенка и продолжается на протяжении всей жизни. Гемопоэз осуществляется в специализированных гемопоэтических тканях: миелоидной (эпифизы трубчатых костей и полости многих губчатых костей) и лимфоидной (тимус, селезенка, лимфатические узлы). В миелоидной ткани образуются эритроциты, лейкоциты, тромбоциты. В лимфоидной ткани идет дальнейшая дифференцировка и созревание лимфоцитов, а также плазматических клеток – основных продуцентов антител.
Постэмбриональный гемопоэз обеспечивает процессы физиологической регенерации крови, то есть её обновление, что компенсирует физиологические процессы разрушения дифференцированных клеток крови.
В условиях нормы функциональная активность органов кроветворения и кроверазрушения строго сбалансирована, что обеспечивает относительное постоянство содержания эритроцитов и других клеток в периферической крови. Разрушение эритроцитов происходит примерно после 120-дневного пребывания их в системной циркуляции при участии тканевых макрофагов селезенки, лимфатических узлов, печени. Гемоглобин, освобождающийся в процессе распада эритроцитов, трансформируется в билирубин в клетках моноцитарно-макрофагальной системы, а затем в гембилирубин (непрямой билирубин), вступая во взаимодействие с белками крови или липопротеидами. Гембилирубин в свою очередь элиминируется из системного кровотока печеночными клетками, где превращается в прямой билирубин (соединение билирубина с глюкуроновой кислотой). Прямой билирубин вместе с желчью поступает в кишечник, постепенно превращается в другие желчные пигменты (стеркобилиноген, уробилиноген) которые, выделяясь с калом и мочой, придают им окраску. При внутриклеточном разрушении эритроцитов основным продуктом, образующимся после распада гемоглобина, является билирубин, а при внутрисосудистом гемолизе большие количества гемоглобина соединяются с α2-гликопротеином-гаптоглобином, который не проникает в мочу.
Основным регулятором эритропоэза является эритропоэтин – гликопротеид, интенсивно вырабатывающийся в условиях гипоксии. При гипоксических состояниях различного генеза концентрация эритропоэтина возрастает в десятки раз по сравнению с нормой. Основным источником синтеза эритропоэтина являются почки (до 90 %), печень (около 10 %), а также макрофаги костного мозга и селезенки. Для эритропоэтина характерен мембранный тип рецепции эритропоэтинчувствительными клетками костного мозга с последующими активацией митоза и дифференцировки клеток, в частности, стимуляцией транспорта железа в эритрокариоцитах, синтеза цепей глобина, ферментов образования гема, синтеза мембранных белков и эритроцитарных антигенов.
Эритропоэз стимулируется под влиянием катехоламинов, глюкокортикоидов, андрогенов, гормонов щитовидной железы, инсулина, плацентарного пролактина, ИЛ-3, ИЛ-6, ИЛ-9, ИЛ-11, КСФ, фолиевой кислоты, витаминов С, В12, железосодержащих препаратов.
Эритропоэз угнетается при повышенной оксигенации тканей, когда снижается образование эритропоэтина, а также под влиянием эстрогенов, глюкагона, ацетилхолина, интерферонов, ФНО-а, ИЛ-1, ИЛ-5, эритроцитарных кейлонов.
После рождения у ребенка в течение нескольких дней отмечают эритроцитоз – содержание эритроцитов составляет 5,5⋅1012/л, в то же время имеется высокое содержание гемоглобина (156-200 г/л). В течение первого года жизни изменяется антигенная структура эритроцитов, возникает прогрессирующее снижение фетального гемоглобина. К концу первого года жизни содержание фетального гемоглобина не превышает 1 %.
Общая характеристика эритроцитов
Эритроциты – самая многочисленная популяция клеток крови, обладающих разнообразными функциями, в частности дыхательной, трофической, детоксицирующей. Количество эритроцитов довольно вариабельно в условиях нормы: так, у женщин оно колеблется от 3,7⋅1012/л до 4,7⋅1012 /л, а у мужчин – от 4,5⋅1012/л до 5,5⋅1012 /л. Сдвиг этих показателей до нижней или верхней границы нормы может быть одним из признаков болезни.
Около 85 % всех эритроцитов имеют форму двояковогнутого диска, то есть являются дискоцитами. Форма эритроцита определяет цепь ауторегулирующих процессов, направленных на поддержание движения крови, её реологических свойств. В условиях патологии появляются эритроциты различной формы, такие состояния именуют пойкилоцитозом. Среди аномальных по форме эритроцитов различают овалоциты, аннулоциты, сфероциты, акантоциты, стоматоциты, щизоциты и другие формы, имеющие в ряде случаев определённое диагностическое значение.
Обычная в условиях нормы форма дискоцита значительно увеличивает площадь диффузии газов, электролитов и других субстратов. Средний диаметр эритроцита (нормоцита) в области краёв составляет 7,5 мкм, а максимальная толщина эритроцита в области краёв составляет 2 мкм. Эритроциты с диаметром от 2 до 6 мкм – микроциты, а с диаметром от 9 до 16 мкм – макроциты. Количество макро- и микроцитов в крови здорового человека в среднем составляет 15–20 %. Резкое увеличение содержания в крови микро- и макроцитов, именуемое анизоцитозом, является одним из признаков нарушения гемопоэза, характерным для анемий, лейкоцитозов, заболеваний инфекционно-аллергической природы. За время свой жизни в периферической крови эритроцит совершает кругооборот более 1 млн. раз, что вызывает развитие механических и метаболических изменений в эритроцитах. Эритроциты обладают пластичностью, то есть способностью к деформации при прохождении через узкие извитые капилляры диаметром 2,5–7,5 мкм. По мере старения их способность к деформации снижается, они застревают в капиллярах красной пульпы селезенки и там разрушаются в процессе фагоцитоза тканевыми макрофагами. Эластичность эритроцитов определяется особенностями структуры белка спектрина, гемоглобина, а также соотношением различных фракций липидов в мембране клеток.
Эритроциты играют важную роль в регуляции кислотно-основного состояния организма, в процессах свертывания крови и фибринолиза за счет адсорбции на их мембране разнообразных ферментных факторов этих систем. Эритроциты являются регуляторами водно-солевого обмена в связи со способностью депонировать воду и минеральные соли при нахождении их в венозной крови. Одной из главных функций эритроцитов является участие в иммунологических реакциях организма за счет наличия в мембранах эритроцитов комплекса полисахаридо-аминокислотных соединений, обладающих свойствами антигенов. Следует помнить, что суммарный объем эритроцитов приблизительно в 160 раз превышает таковой лейкоцитов и тромбоцитов, а потому кровь можно рассматривать как двухфазную систему, представляющую собой взвесь (суспензию) эритроцитов в плазме. При этом эритроцит подвергается в токе крови действию напряжения сдвига – оно более значительно у края и направлено в сторону стенки и менее выражено в центре сосуда. Разница действующих векторов силы у разных краёв обеспечивает вращательное движение эритроцитов в текущей жидкости, что при столкновении с тромбоцитами, имеющими меньший размер и худшую деформируемость по сравнению с эритроцитами, приводит к отбрасыванию кровяных пластинок к стенке сосуда. Благодаря этому пристеночный слой оказывается обогащенным тромбоцитами. Указанный эффект обусловлен величиной гематокрита, размером эритроцитов и тромбоцитов и ригидностью их мембран. Увеличение любого из указанных параметров сопровождается усилением передвижения кровяных пластинок к стенке сосуда, а при наличии повреждения эндотелия – адгезией к субэндотелию.
Буферные свойства эритроцитов. Как известно, на единицу объема эритроцит связывает в 60 раз большее количество О2, чем плазма крови. О2 хорошо растворим в воде, поэтому диффузия его в растворе происходит очень быстро. Связывание О2 с эритроцитами определяется парциальным давлением газа в плазме и сорбционными свойствами Нв. В капиллярах легких, где давление О2 высоко (рО2 = 133 гПа), высоко и сродство Нв к О2, что обеспечивает трансмембранный перенос газа и связывание его с гемоглобином. В капиллярах тканей, где рО2 равно 40-50 гПа, сродство Нв к О2 резко снижается. При этом происходит выход кислорода из эритроцитов.
Транспорт СО2 через мембрану эритроцита также осуществляется за счет диффузного давления (в капиллярах легких рСО2 = 53гПа, а в капиллярах тканей – 61 гПа). Диффузия СО2 в растворах происходит примерно в 20 раз быстрее, чем О2.
Высокая скорость равновесия содержания СО2 в системе эритроцит – плазма крови обеспечивается наличием в эритроцитах особого фермента – карбоангидразы, катализирующего реакции трансформации СО2 и Н2О в углекислоту (Н2СО3), а также мощными системами трансмембранного обмена анионами.
При понижении концентрации СО2 в эритроцитах возникает отрицательный заряд Нв, что приводит к уменьшению содержания внутриклеточной воды, а при увеличении содержания СО2 в эритроцитах – они набухают.
Известно, что осмотическое давление в эритроцитах несколько выше, чем в плазме крови, что связанно с высокой внутриклеточной концентрацией белков по сравнению с плазмой крови. При этом содержание низкомолекулярных осмотически активных веществ (ионов натрия) в эритроцитах значительно меньше, чем в плазме крови. Величина осмотического давления в эритроцитах обеспечивает достаточный или нормальный тургор этих клеток. Осмотическое давление плазмы и эритроцитов в условиях нормы находится в динамическом равновесии, что обуславливает стабильность структуры эритроцитов.
При помещении эритроцитов в коллоидно-осмотическую среду с более низким осмотическим давлением (гипотонические растворы) может наступить осмотический или коллоидно-осмотический гемолиз. Последний обусловлен тем, что вода поступает в эритроциты до того момента, пока не разрывается мембрана и гемоглобин выходит в окружающую среду.
В умеренногипотонической среде эритроциты приобретают сферическую форму, их называют в связи с этим сфероцитами. Способность эритроцитов сохранять свою структуру при развитии гипоосмотических состояний или в гипоосмотической среде получила название осмотической устойчивости, или резистентности эритроцитов. Верхняя граница резистентности или максимальная устойчивость эритроцитов соответствует примерно 0,5–0,4 % раствора хлорида натрия.
При помещении эритроцитов в гипертоническую среду происходит их сморщивание в связи с потерей воды и уменьшением объема.
Эритроцитам свойственна способность к оседанию. Удельная масса цельной крови в норме для взрослого составляет в среднем 1,05–1,06. Удельная масса эритроцитов (1,085–1,096) выше, чем плазмы крови (1,02–1,027), поэтому эритроциты в пробирке с кровью, лишенной возможности свертываться, способны медленно оседать на дно. Скорость оседания эритроцитов в значительной мере определяется белковым составом плазмы крови, в частности уровнем мелкодисперсных белков-альбуминов. В связи с этим важная роль в обеспечении величины СОЭ отводится соотношению альбуминово-глобулиновых фракций белков крови. СОЭ у мужчин в среднем составляет 1–10 мм/ч, у небеременных женщин 2-15 мм/ч. При некоторых патологических процессах и заболеваниях, а также во второй половине беременности СОЭ повышается, так как увеличивается содержание в крови грубодисперсных белков глобулиновой фракции, получивших название аггломеринов, а также за счет усиления образования фибриногена.
При замедлении скорости кровотока и повышении вязкости крови эритроциты проявляют способность к агрегации. Вначале агрегация носит обратимый характер, при этом образуются ложные агрегаты, или монетные столбики. В случае быстрого восстановления кровотока они распадаются на полноценные клетки с сохраненной мембраной и внутриклеточной структурой.
Пластичность или деформируемость – это способность эритроцитов к обратимой деформации при прохождении через узкие извитые капилляры, микропоры. Данное свойство определяется особенностями структуры мембраны эритроциты, наличием в ней особого белка спектрина.
Таким образом, основными физиологическими и физико-химическими свойствами эритроцитов являются следующие:
– способность к оседанию;
– способность к агрегации;
– деструкция после определенного периода циркуляции в кровотоке.
1. Дыхательная функция заключается в захвате и переносе кислорода к тканям и экскреции СО2 из организма. Это обеспечивается содержащимся в эритроцитах белком гемоглобином. Гемоглобин – сложный белок состоит из групп гема и белкового остатка – глобина. Содержание гемоглобина у мужчин составляет 130-160 г/л, у женщин 120-140 г/л.
2. Трофическая функция эритроцитов связана с их способностью транспортировать аминокислоты, нуклеотиды, пептиды к различным органам и тканям, способствуя обеспечению репаративных процессов. В ряде случаев эту функцию называют транспортной.
3. Детоксиксицирующая функция эритроцитов обусловлена их способностью адсорбировать токсические продукты эндогенного или экзогенного происхождения и частично инактивировать их.
4. Участие в процессах свертывания крови за счет адсорбции на их мембране плазменных факторов свертывания крови
5. Участие в регуляции кислотно-основного состояния организма (буферная функция) за счет гемоглобина обеспечивающего до 70 % буферной ёмкости крови.
6.– Ферментативная функция связана с наличием в эритроцитах большого количества ферментов, в частности карбоангидразы, метгемоглобинредуктазы, ферментов гликолиза.


